|
Дипломная работа: Взаимодействие микроорганизмов с высшими растениями
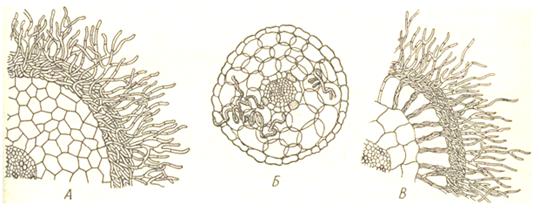
Рис. 6. Поперечные срезы микориз древесных пород. Микоризы: А – эктотрофная (береза); Б – эндотрофная (клен); В – эктоэндотрофная (дуб). Гифы гриба проходят между клетками эпидермиса, образуя сеть. Такая микориза характерна для древесных растений и редко встречается у травянистых (4, 221). Провести четкую границу между эндотрофной и экзотрофной мико-ризами трудно. Переходом между ними являются эктоэндотрофная микориза, распространенная более часто, чем чисто эктотрофная. Грибные гифы при такой микоризе густо оплетают корень снаружи и в то же время дают обильные ветви, проникающие в его коровую паренхиму. Мицелий идет отчасти межклетно, отчасти внутриклетно, образуя в клетках клубки, гиф или разветвления, напоминающие гаустории. Клетки корня при этом остаются живыми (см. Рис.6). Такая микориза встречается у большинства древесных пород: дуб, ель, сосна, лиственница и другие (17, 233). Микоризу образует большинство растений (за исключением водных), как древесных, так и травянистых (особенно многолетних). Травянистые растения вступают в микоризный симбиоз с микроскопическими грибами в основном из класса несовершенных грибов (Deuteromycetes), отчасти из класса зигомицетов (Zygomycetes) с мицелием, лишенным перегородок (неклеточным) и отчасти из класса сумчатых грибов (Ascomycetes). Грибы родов элафомицес (Elaphomyces) и трюфель (Tuber) образуют микоризу с буком, дубом и другими деревьями. Но большинство древесных пород образуют микоризу с грибницей шляпочных грибов – макромицетов из класса базидиальных (Basidiomycetes) и группы порядков гименомицетов. В гименомицетной эктоэндотрофной микоризе древесных пород гриб оплетает корень снаружи и частично проникает внутрь. Здесь он получает от корня углеводное питание, так как сам, будучи гетеротрофом, не может синтезировать органические вещества. Наружные свободные гифы гриба широко расходятся в почве от корня, заменяя последнему корневые волоски. Эти свободные гифы получают из почвы воду, минеральные соли, а также растворимые органические вещества (главным образом азотистые). Часть этих веществ поступает в корень, а часть используется самим грибом на построение грибницы и плодовых тел. В клетках корня частично перевариваются внедрившиеся гифы и служат дополнительным источником органических веществ. Почва леса, особенно в прикорневой зоне деревьев, пронизана грибницей микоризных грибов, а на поверхности почвы появляются многочисленные плодовые тела. Это подберезовик (Leccinum scabrum), подосиновик (Leccinum aurantiacum) и многие другие шляпочные грибы, встречающиеся только в лесу. Для этих грибов такой симбиоз обязателен. Если их грибница и может развиваться без участия корней дерева, то плодовые тела в этом случае обычно не образуются. Белый гриб образует микоризу со многими породами деревьев: березой, дубом, сосной, елью. Некоторые виды грибов – только с одной определенной породой. Так лиственничный масленок образует микоризу с лиственницей. Для деревьев симбиоз с грибами тоже имеет значение: опыты на лесных полосах показали, что без микоризы деревья развиваются хуже, отстают в росте, они более ослаблены, подвержены заболеваниям. В случае эндотрофных микориз взаимоотношения гриба и высшего растения еще более сложные. В связи с малым контактом гиф микоризного гриба с почвой таким путем в корень поступает относительно небольшое количество воды, минеральных и азотистых веществ. В этом случае значение для высшего растения, вероятно, приобретают вырабатываемые грибом биологически активные вещества типа витаминов. Отчасти гриб снабжает высшее растение азотистыми веществами, так как часть гиф гриба, находящаяся в клетках корня, переваривается ими. Гриб получает углеводы, а в случае микоризы орхидных гриб сам отдает углеводы (сахар) высшему растению (17, 234). Нужно отметить, что микоризный симбиоз – явление очень сложное, не укладывающееся в какую-то определенную схему. Эти взаимоотношения часто зависят от окружающих условий. Некоторые микроорганизмы ризосферы постепенно переходят в состав эпифитной микрофлоры. Особенно оживился этот вопрос в связи с работами немецкого ученого Е. Либберта (1966), который выступил с утверждением, что гормон роста высших растений гетероауксин синтезируется не растением, а эпифитной микрофлорой. Работы В.И. Кефели (1969, 1971) показали, что в стерильных условия капуста образует гетероауксин из L – триптофана. А.А. Тарасенко (1972) установила, что эпифитная микрофлора влияет положительно на рост и обмен веществ проростков кукурузы (19, 142). Микроорганизмы, развивающиеся на поверхности стеблей или листьев растений, получили название эпифитной микрофлоры. Микроорганизмы-эпифиты вынуждены довольствоваться минимальными источниками питательного субстрата, представленного выделениями растительных тканей и веществами-загрязнителями (пылью). Поэтому состав эпифитной микрофлоры весьма специфичен. Нередко 80% общего количества эпифитов составляют бактерии Erwinia herbicola. Второе место по численности занимают различные грибы (Penicillum, Mucor, Fusarium и другие). На поверхности многих тропических растений обнаружены азотфиксирующие бактерии рода Beiyerckia, поставляющие азот непосредственно в лист. Разнообразная и обильная микрофлора находится на поверхности семян. Так, на 1г зерна ржи приходится не менее 2.500 тыс. микробных клеток, пшеницы – 1.500 тыс., риса – 250 тыс. В состав микрофлоры зерна обязательно входят неспорообразующие бактерии Pseudomonas, Arthrobacter и Flavobacterium, дрожжи Candida, Rhodotorula, Criptococcus, а также грибы Penicilium, Aspergillis, Alternaria, Cladosporium, Mucor и другие. Развитие микроорганизмов на поверхности зерна в значительной мере зависит от влажности и температуры. Установлено, что при температуре 15 – 20°С и влажности 14,5 – 15% на зерне пшеницы начинается развитие грибов, а при влажности 17,5 – 18% - бактерий. Общая численность эпифитных микроорганизмов резко возрастает при повышении влажности воздуха и усиленном выделении продуктов обмена растительными тканями. При жизни растения эпифитная микрофлора не оказывает на него вредного влияния. Наоборот, питаясь продуктами выделений растений, она способствует освобождению тканей от продуктов собственного обмена (15, 259). Эпифитная микрофлора образует определенный биологический барьер, препятствующий заражению растительных тканей фитопатогенными микробами (26, 83). Таким образом, симбиотические взаимоотношения микроорганизмов с высшими растениями разнообразны. При таком контакте пользу получают оба организма, не нанося друг другу вреда. 1.3 Влияние фитопатогенных микроорганизмов на высшие растения Первые сведения о патологических изменениях в тканях растений были сделаны М. С. Ворониным (1867) при изучении образования клубеньков на корнях люпина. Это явление, хотя и симбиотическое, дало возможность обратить внимание на паразитический характер жизни микроорганизмов в растениях. Практически во всех группах микроорганизмов имеются возбудители болезней растений (см. Рис. 7).
грибы бактерии вирусы актиномицеты Рис. 7. Группы фитопатогенных микроорганизмов. Первое место среди фитопатогенных микробов принадлежит грибам, второе место занимают бактерии и вирусы, и лишь небольшой процент болезней растений вызывают актиномицеты. Растения больше поражаются грибами, чем бактериями. Это связано с более кислой средой тканей растений, которая благоприятствует развитию больше грибов, чем бактерий. Тем не менее, известно довольно много бактериальных болезней растений (бактериозами) (10, 221). Источники заражения фитопатогенными микроорганизмами различны. Одним из важнейших источников заражения являются семена. Попадая внутрь или на поверхность семян, фитопатогенные микроорганизмы находят подходящее место для перезимовки. При прорастании семян они могут заражать всходы, а затем по проводящим сосудам передвигаться в растения и заражать взрослые растения в период вегетации. Кроме того, больные семена могут служить источником распространения инфекции. Заболевание могут распространять зеленые растения, в которых микробы хорошо сохраняются и переносятся в новые районы вместе с зараженными растениями (черенки, окулировочные материалы — глазки). Одним из основных источников заражения бактериозами являются остатки больных растений. Особенно долго и хорошо фитопатогенные микроорганизмы сохраняются в деревянистых частях растений. Некоторые виды насекомых также могут являться источником первичной инфекции. Большую опасность в распространении бактериозов представляют капельки дождя с мелкими частицами остатков больных растений, которые ветром и воздушными течениями разносятся на далекие расстояния. Переносить фитопатогенные микроорганизмы может также и вода — поливная, вода рек и других источников. Человек может распространять заболевания растений на большие расстояния при перевозке семян и посадочного материала, а также при обработке растений, уходе за ними в период вегетации (16, 399). Проникший в ткани растения паразит выделяет различные вещества, комплекс которых получил название токсинов. В состав токсинов входят ферменты, могут входить некоторые органические кислоты и амины, специфические полисахариды и другие разнообразные соединения. Выделяя токсины, паразит убивает клетки растения-хозяина, питаясь продуктами разложения этих мертвых клеток. Иными словами, в ряде случаев имеется настоящий внутренний сапрофитизм. Как установили исследования К. Т. Сухорукова и его учеников, гриб, вызывающий заболевание, известное под названием вилта хлопчатника, действительно убивает своими выделениями клетки растения-хозяина и затем их использует для своего питания. Паразитические формы грибов и бактерий возникли из сапрофитических форм, и явление эндосапрофитизма свидетельствует об этом. Такое же явление можно отметить для гриба Botrytis hyssoidea. Он проникает через отмершие ткани как сапрофит в верхушку луковицы, затем становится паразитом, развиваясь на живых, растущих тканях, убивая их, продолжает на них своё развитие как сапрофит (10, 221). Особенно токсичными выделениями, убивающими клетки растения-хозяина, отличаются факультативные паразиты, которые могут жить как сапрофиты. Поселяясь на живом растении, они предварительно его убивают. В большинстве случаев такие факультативные паразиты являются мало специализированными. Облигатные (обязательные) паразиты обычно строго специализированы в отношении растения-хозяина. Многочисленные примеры строгой специализации дают ржавчинные и головневые грибы-паразиты (10, 221). Взаимодействие между растением-хозяином и паразитом проходит ряд этапов. Можно различить четыре фазы в процессе поражения растения паразитом (Б.А. Рубин, 1964). В первой фазе паразит (обычно спора гриба или бактерии), попадая на покровные ткани растения, начинает прорастать за счет веществ, вымываемых из тканей растения росой, дождем, поливом. Чем больше питательных веществ вымывается из растений, тем легче происходит заражение. Отмечено, что у устойчивых к гоммозу (Bact. Malvacearum) сортов хлопчатника количество вымытых веществ в два раза меньше, чем у неустойчивых. Выделяемые растением питательные вещества вызывают хемотропическое или хемотаксическое движение паразита к тканям растения. Вторая фаза заключается в проникновении паразита внутрь растения. Большое значение во втором этапе имеют многие анатомические свойства растений, в частности толщина кутикулы и оболочки и т.д. Паразит или разрушает механически оболочку клетки, чему способствует свойственное большинству паразитических грибов высокое осмотическое давление (50 и более атмосфер), или чаще происходит растворение кутикулы, оболочки и особенно срединных пластинок оболочек внедряющимся паразитом. Многие паразиты проникают сквозь устьица. В ответ на внедрение многие организмы защищаются образованием специализированных клеток. При этом всегда возрастает интенсивность дыхания (10, 222). Третья фаза представляется центральной для явлений иммунитета, так как здесь происходит основное физиологическое взаимодействие между растением-хозяином и паразитом. Последний мобилизует все имеющиеся у него средства нападения, а растение-хозяин – все имеющиеся у него средства защиты. Растение реагирует неодинаково на факультативного неспециализированного паразита и на облигатного. Факультативный паразит, например, гриб Botrytis, выделяет в значительном количестве токсины, убивающие клетки растений. Устойчивые растения подавляют действие токсинов, в результате чего не происходит большого некроза тканей. У неустойчивых видов наблюдается полная деформация и некроз тканей. Облигатные паразиты действуют гораздо медленнее на растение-хозяина, и это позволяет им завершить в организме цикл своего развития. Растение реагирует на внедрение облигатного паразита образованием и накоплением в протоплазме темно-окрашенных продуктов окисления – фенолов, пропитывающих в дальнейшем оболочку клеток. Эти изменения вызывают гибель клетки и часто сопровождаются гибелью паразита. Клевер, пораженный грибком Erysiphe polygonii, дает у восприимчивого сорта картину более или менее мирного сосуществования гриба и паразита, заканчивающегося истощением растения-хозяина и спороношением паразита. У устойчивого к этому заболеванию сорта клевера в месте инфекции наблюдается резкое потемнение, некроз части клеток и сильный подъем окислительных процессов. Изменение окислительных процессов под влиянием паразита является ответной защитной реакцией организма хозяина. При этом снижается активность гидролитических ферментов микроорганизмов и растения-хозяина, что препятствует переводу питательных веществ в доступную для паразитов форму. Высокая интенсивность окислительных процессов приводит к обезвреживанию токсинов, а также способствуют синтезу веществ (суберин), препятствующий распространению паразитов. Окисление фенолов приводит к образованию хинонов, которые, пропитывая вещества клетки, делают их недоступными для питания паразитов. Четвертую фазу можно считать заключительным периодом взаимодействия обоих организмов. Наблюдаемая здесь картина зависит от степени устойчивости растения-хозяина. Иммунные формы в это время полностью ликвидируют инфекцию, и все процессы приходят в норму. У не иммунных форм в этот период появляется ясно выраженное заболевание с полным нарушением обмена веществ и распадом тканей (10, 223). Наиболее распространенные инфекционные заболевания, вызываемые: - бактериями – это загнивание, опухали, рак, бактериозы, ожоги ветвей, листьев, плодов; - грибами – головня, ржавчина, спорынья, мучнистая роса (см. Приложение 3); - вирусами – мозаичные болезни, морщинистость или карликовость листьев (13, 31). Устойчивость растений против инфекционных заболеваний называется иммунитетом. Есть немало растений, которые не поражаются той или иной инфекционной болезнью. Различают иммунитет абсолютный – полную устойчивость к данному заболеванию (например, у пшеницы к пыльной головне овса) и относительный – частичную поражаемость растений в зависимости от условий окружающей среды (11, 219). Большой вклад в учение об иммунитете внесли исследования Н.И. Вавилова. Он установил ряд анатомо-морфологических и физиологических особенностей (10, 119). К анатомо-морфологическим особенностям относится наличие кутикулы и пробки, покрывающей часто органы растения, а также состоящих из целлюлозы оболочек, окружающих каждую клетку. Всё это препятствует быстрому распространению болезни по растению. Поэтому многие паразиты проникают в растение сквозь устьичные щели, как это, например, отмечено для картофельного гриба (Phitophtora infestans). Паразит, проникший в растительный организм, вызывает у него ответную реакцию. Больное растение отличается от здорового целым рядом свойств. В работе А.Л. Курсанова было показано значительное усиление дыхания и транспирации под влиянием головневого гриба, поражающего злаки. Несколько повышался фотосинтез, но это повышение было неустойчиво и не компенсировало траты вещества за счет усиленного дыхания. Таким образом, внедрение паразита в ткани высшего растения вызывает физиологические изменения. Увеличивается проницаемость цитоплазмы и нарушается водный обмен, приводящий к повышению транспирации, возрастает активность гидролитических ферментов, что влечёт за собой распад ряда сложных соединений клетки (белков, крахмала) и их превращение в доступные паразиту более простые формы, разрушающие хлорофилл и частично пластиды, нарушающие весь обмен веществ (дыхание, фотосинтез, ферментативная деятельность). Л.И. Курсанов (1933) наблюдал отрицательное влияние гриба Chrysomyxa piola на грушанку Piola rotundifolia в первый год заражения. На второй год, когда у гриба наступало спороношение, и он нуждался в значительном количестве питательных веществ, наблюдалось резкое нарушение функций пораженного грибом растения (10, 220). В литературе отмечают деструктивных паразитов и сбалансированных. Первые вызывают полную гибель растения, а вторые могут жить годами в симбиозе с растением-хозяином. Факультативные паразиты, грибы из рода Botrytis, представляют собой деструктивных паразитов, а вышеприведённый пример с грибом Chrysomyxa – сбалансированных. Однако сбалансированный паразитизм у Chrysomyxa на второй год жизни переходит в деструктивный. Помимо чисто анатомо-морфологических и физиологических изменений, растения обладают химическими средствами защиты, связанными с накоплением в клетках тканей различных химических веществ – гликозидов, алкалоидов, фенольных соединений, губительно действующих на фитопатогенных микроорганизмов. Такими веществами являются фитонциды и фитоалексины (10, 220). В иммунитете большую роль играют фитонциды (от греч. «фитон» – растение и лат. «цидере» – убивать), открытые Б.П. Токиным в 1928г. Учёный установил их наличие у многих высших растений. Фитонциды могут выделяться растением в газообразном, а также в растворенном в воде виде. Особенно богаты фитонцидами чеснок, лук, тысячелистник, подорожник, берёза и некоторые другие растения. Химическая природа фитонцидов неодинакова. У одних растений это органические кислоты, у других – эфирные масла, алкалоиды или аминокислоты. Они останавливают рост микроорганизмов, повреждают и убивают их (11, 144). Фитонциды черёмухи и лука быстро убивают картофельный гриб (Phitophtora infestans) (10, 221). Многие фитонциды действуют обычно только в течение небольшого промежутка времени после их выделения из растений. По-видимому, открытые цепи соединений, обладающих фитонцидным действием, замыкаются в кольца и теряют свою активность. Фитоалексины (от слов «фитон» - растение и «алексо» – отражение атаки) были открыты в 1940г. Они относятся к своеобразным антибиотикам и образуются только высшими растениями в ответ на заражение их фитопатогенными микроорганизмами. Фитоалексины отличаются от фитонцидов и других антибиотиков тем, что являются производными двух систем – растения-хозяина и паразита. Метаболиты паразита являются индукторами, вызывающими отклонения в обмене веществ у растения-хозяина. Последнее образует специфические для данного вида фитоалексины. Образование фитоалексинов идёт в живых клетках растения, но максимальные их количества находятся в некротической ткани. В этом проявляется реакция сверхчувствительности растения, при которой за счёт гибели части клеток организм как единое целое сохраняется. В некоторых случаях растение образует два фитоалексина, что затрудняет проникновение в него паразита. Так, например, у картофеля на инфекцию картофельного гриба (Phytoptora infestans) образуются токсичные фитоалексины решитин и любимин. Все фитоалексины представляют собой низкомолекулярные соединения, а вещества, выделяемые паразитом, являются соединениями белковой природы. Многие стороны учения о фитоалексинах разработаны Л.В. Метлицким и О.Л. Озерецковской (1973 г.) (10, 223). Таким образом, большинство фитопатогенных микроорганизмов активно синтезируют гидролитические ферменты (пектиназы, целлюлазы, протеазы и др.), вызывающие разрушение растительных тканей и клеточных оболочек, что проводит к проникновению возбудителя болезни внутрь клетки. Проникнув в клетку, фитопатогенные микробы нарушают нормальный ход физиологических процессов, прежде всего фотосинтеза и дыхания. Токсины, выделяемые возбудителем болезни, инактивируют ферменты растительной клетки, что в конечном счете приводит её к гибели. Растения защищаются различными способами от фитопатогенных микроорганизмов. Выводы по главе I Обобщив выше сказанное, сделаем следующие выводы: 1) Взаимодействие микроорганизмов с высшими растениями – это взаимная связь организмов величиной от 50 до 500 мкм с автотрофными формами, имеющими расчленение на листья, стебли и корни. 2) Между высшими растениями и микроорганизмами существуют различные симбиотические взаимоотношения, приносящие пользу взаимодействующим организмам. К симбионтам высших растений относятся ризосферные и эпифитные микроорганизмы, клубеньковые бактерии, грибы-микоризообразователи. 3) Практически во всех группах микроорганизмов имеются возбудители заболеваний растений. Различают 4 фазы поражения растения паразитом. Проникнув в клетку, фитопатогенные микробы нарушают нормальный ход физиологических процессов в растении. 4) Устойчивость растений против инфекционных заболеваний называется иммунитетом. Существуют анатомо-морфологические и физиологические особенности растений, способствующие их невосприимчивости к инфекциям. Особое место занимают фитонциды и фитоалексины. Глава II. Место и роль знаний о взаимодействии микроорганизмов с высшими растениями в школьном курсе биологии 2.1 Изучение взаимодействия микроорганизмов с высшими растениями на уроках биологии Социальный заказ школе, учитывающей достижения биологической науки и изменения в окружающем мире, предъявляет требования к школьному биологическому образованию, к повышению биологической грамотности подрастающего поколения. В обучении мы должны исходить из того, что биологическая грамотность становится социально необходимой, поскольку основной объект биологии – жизнь, а сейчас остро стоят вопросы, как выжить, как обращаться с природой, чтобы не нанести ей вреда, проявить заботу не только о себе, но и об окружающей среде (41, 53). Биологическое образование должно обеспечить выпускникам высокую биологическую и экологическую грамотность. Решить эту задачу можно лишь на основе преемственного развития понятий, законов, теорий, идей, являющихся фундаментом для практической деятельности учащихся, формирования их научного мировоззрения (32, 57). При анализе «Обязательного минимума содержания основного общего образования по биологии» мы выяснили, что часть материала нашей работы является обязательной для изучения. Сюда входит: бактерии и грибы – возбудители заболеваний растений, биологические основы выращивания культурных растений (27, 29). Следует учитывать, что в минимум включены не все вопросы содержания. Это означает, что минимум ориентирует учителя на более широкое и глубокое раскрытие содержания каждой темы. В системе обучения биологии ведущая роль принадлежит уроку, который был и остается основной формой учебно-воспитательной работы в школе. На уроках биологии учитель дает теоретические знания, формирует у учащихся важнейшие биологические понятия. Наиболее часто встречающимся элементом урока является изучение нового материала. Учитель вооружает учащихся системой биологических знаний, переводит их в умения и навыки через организацию определенных программой лабораторных и практических работ, формирует общеучебные умения (30, 8). При разработке поурочного планирования учитель решает, какой дополнительный материал можно использовать, чтобы заинтересовать учащихся. При этом учитывается важность и полезность информации. В дальнейшем этот материал должен быть востребован учителем. |
|
|||||||||||||||||||||||||||||
|
|||||
Рефераты бесплатно, реферат бесплатно, сочинения, курсовые работы, реферат, доклады, рефераты, рефераты скачать, рефераты на тему, курсовые, дипломы, научные работы и многое другое. |
||
При использовании материалов - ссылка на сайт обязательна. |
||