|
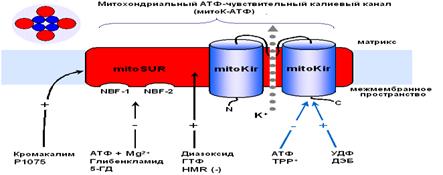
Дипломная работа: Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданиюМитоКАТФ, также как и клеточный, является, по всей вероятности, гетеромультимером, состоящим из калиевого канала, белка с молекулярной масой 55 кДа, который имеет выпрямляющие свойства и который, по аналогии с цитоплазматическим каналом, был назван митоKIR (от англ. Inward rectifying) [Mironova et al., 2004] и рецептора, чувствительного к сульфонилмочевинам и поэтому названного митоSUR (от англ. sulphonylurea) (Рис. 3) [Mironova et al., 1999; Bajgar et al., 2001]. Кi для АТФ на реконструированном в БЛМ митоKIR выше, чем в интактных МХ и Mg2+ для этого ингибирования не требуется. Тогда как для АТФ-ингибирования канала, локализованного в интактных МХ, присутствие магния было необходимо [Mironova et al., 2004]. Те же различия были обнаружены при сравнительном изучении влияния АТФ и сульфонилмочевин на цитоKIR и целый цитоКАТФ [Tucker et al., 1997]. Известно, что основной участок связывания АТФ в цитоКАТФ локализован на канальной субъединице. Регуляторная субъединица повышает сродство канальной субъединицы к АТФ и обеспечивает чувствительность целого канала к активаторам и ингибиторам [Tucker et al., 1997].
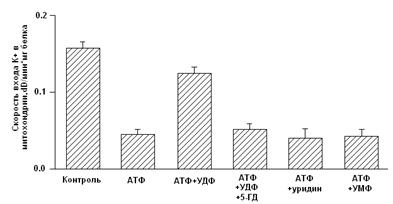
Рис.3. Рабочая модель структуры МХ АТФ-зависимого К+ канала [Mironova et al., 2004] Приведенные выше результаты исследований митоКАТФ подтверждают предположение о том, что белок с м.м. 55 кДа является канальной субъединицей целого митоКАТФ. Функцию регуляторной субъединицы выполняет, вероятно, белок с м.м. 63 кДа, связывающийся с меченым глибенкламидом [Bajgar et al., 2001]. В то же время, результаты исследований, проведенных на МХ сердца, с использованием антител на Kir6.1 и Kir6.2, а также на SUR1 и SUR2, показали, что в МХ кардиомиоцитов присутствуют Kir6.1 и Kir6.2, но нет ни SUR1, ни SUR2. Однако в них обнаруживается SUR2-подобный белок, который, возможно, является сплайсированным вариантом SUR2 [Lacza et al., 2003 (I)]. При исследовании митоКАТФ мозга, близкого по структуре к цитоКАТФ [Lacza et al., 2003 (II)], с использованием АТ к KIR6.1 и KIR6.2, а также SUR1 и SUR2 субъединицам цитоКАТФ было показано, что очищенные МХ мозга содержат преимущественно KIR6.1 и, в меньшей степени, KIR6.2 и не содержат SUR1 и SUR2. Согласно выводам авторов, в формировании митоКАТФ мозга участвует укороченный вариант SUR2 или белок, подобный по структуре SUR2. Кроме того, показано, что МХ мозга содержат в 8 раз больше KIR6.1 и SUR2-подобного белка, по сравнению с общим белком ткани мозга, что подтверждает локализацию этих белков в МХ [Lacza et al., 2003 (II)]. Тот факт, что м.м. митоKIR мозга несколько ниже (50 кДа), чем у митоKIR печени [Mironova et al., 1999; 2004], предполагает наличие тканевой специфичности в структуре каналов. Было показано, что при встраивании в липосомы или липидные бислойные мембраны комплекса митохондриальных белков, состоящего из АТФ связывающего кассетного белка-1 (mАВС1 от англ. Mitochondrial binding casset), АТФ-азы, сукцинатдегидрогеназы, аденилаттранслоказы и фосфатного переносчика, в мембране образуются чувствительные к АТФ и глибенкламиду каналы [Ardehali et al., 2004]. Это послужило основой для другого варианта модели субъединичной организации митоКАТФ [Ardehali and O'Rourke, 2005]. Однако нельзя исключить наличие в этом белковом комплексе других митохондриальных белков, включая митоKIR, тем более что белок с м.м. 55 кДа в нём присутствует. Поcкольку этот мультикомплекс в липосомах индуцировал открытие неспецифической Са2+-зависимой поры в присутствие ионов Са2+ [Ardehali et al., 2004], это говорит о возможном загрязнении исследуемого комплекса порином и другими белками. Было показано, что один из белков этого комплекса, а именно mАВС1, играет важную роль в защите клеток миокарда от окислительного стресса [Ardehali et al., 2005]. Однако кардиопротекторное действие этого белка не снималось ингибиторами митоКАТФ - глибенкламидом и 5-НD. Поэтому, по мнению авторов, пока не получено достаточно доказательств для утверждения того, что mАВС1, имеющий, также как и митоКIR, м.м. 55 кДа, является частью митоКАТФ. Таким образом, вопрос структуры митоКАТФ по-прежнему остается открытым. В связи с чем, в представленной работе была предпринята попытка определить относится ли белок с м.м. 55 кДа, выделяемый из внутренней мембраны МХ методом водно-этанольной экстракции, к системе АТФ-зависимого транспорта К+. Также, был проведен анализ гомологии структуры исследуемого белка последовательностям известных белков. Глава 2. Модуляторы 2.1 Модуляторы митоКАТФ канала 2.1.1 Метаболические модуляторы митоКАТФ канала В связи с выраженным кардиопротекторным действием активаторов митоКАТФ в настоящее время во многих лабораториях ведется поиск новых синтетических активаторов канала, которые могут быть потенциальными кардиопротекторами [Ockaili et al., 2002; Kopustinskene et al., 2001]. В нашей лаборатории помимо синтетического донора электронов ДЕБ, который селективно активировал митоКАТФ [Mironova et al., 1999; Mironova et al., 2004], был обнаружен ряд природных, то есть метаболических активаторов этого канала. Дифосфонуклеотиды Показано, что дифосфонуклеотиды (АДФ и ГДФ) активируют реконструированную в БЛМ канальную субъединицу (Рис. 2) [Grigoriev et al., 1999; Mironova et al., 1999]. Данные по активирующему действию ГДФ в дальнейшем были подтверждены и другими исследователями [Bednarczyk et al., 2005]. Недавно в нашей лаборатории было показано, что в ряду дифосфонуклеотидов наиболее эффективным является уридиндифосфат (УДФ) [Mironova et al., 2004]. В МХ УДФ (~20 мкМ) активирует митоКАТФ, и эта активация снимается глибенкламидом и 5-ГД (Рис. 4) [Mironova et al., 2004]. Концентрации УДФ, неоходимые для активации реконструированного в БЛМ митоKIR и целого канала в нативных МХ, одинаковы. Следовательно, участок связывания УДФ локализуется на канальной субъединице. Для активации цитоКАТФ нужны значительно большие концентрации этого препарата [Alekseev et al., 1998].
Рисунок 4. Активация УДФ целого митоКАТФ канала в нативных МХ [Mironova et al., 2004] Ранее на моделях тотальной и регионарной ишемии изолированных перфузируемых сердец крыс было показано, что уридин и УМФ проявляют антиаритмическую активность в отношении реперфузионных аритмий, предупреждают развитие миокардиального станинга при постишемической реперфузии, препятствуя депрессии сократительной функции миокарда и предотвращая развитие контрактуры [Eliseev et al., 1989]. В представленной работе была предпринята попытка определить, обладают ли уридиновые нуклеотиды кардиопротекторным действием и связано ли их влияние с активацией цито- и митоКАТФ каналов на модели искусственного острого инфаркта миокарда. 2.2 Функциональная роль митоКАТФ Являясь одним из компонентов калиевого цикла МХ, митоКАТФ участвует в поддержании объема МХ матрикса и мембранного потенциала [Garlid, 1998]. В лаборатории проф. Мироновой показано, что активация митоКАТФ играет важную роль при выходе животных из состояния зимней спячки [Fedotcheva et al., 1985; Миронова и др., 1986]. МитоКАТФ участвует также в ингибировании процесса апоптоза клетки, блокируя высвобождение цитохрома С, предотвращая снижение мембранного потенциала и чрезмерное увеличение концентрации Ca2+, сохраняя, таким образом, функции МХ, что было показано на b-клетках поджелудочной железы [Akao et al., 2001], а затем и на церебральных нейронах [Teshima et al., 2003]. Кроме того, митоКАТФ является посредником в процессе регуляции транскрипции некоторых генов. Его активация вызывает небольшое увеличение продукции активных форм кислорода (АФК) [Chandel et al., 1998]. При патологии активация митоКАТФ частично востанавливает мембранный потенциал, стимулируя дальнейший перенос протонов, ингибируя накопление внутримитохондриального кальция и предотвращая распад энергетических фосфатов путем создания оптимального для синтеза АТФ электрохимического градиента [Szewczyk, 1996; Xu et al., 2001]. Активация митоКАТФ способствует разобщению электронного транспорта, ускоряя, таким образом, дыхание, что приводит к окислению проксимальных переносчиков электронов дыхательной цепи [Liu et al., 1998]. Согласно данным Гарлида, активация канала приводит к набуханию МХ, что способствует поддержанию архитектуры внутренней мембраны и сохранению активности дыхательной цепи. Это позволяет поддерживать уровень клеточного АТФ и, таким образом, способствует восстановлению функций миокарда во время реперфузии [Garlid, 2000]. Как было показано ранее, набухание матрикса увеличивает скорость окислительного метаболизма, активирует как окисление жирных кислот, так и дыхание и синтез АТФ [Halestrap, 1989]. 2.2.1 Активация митоКАТФ в развитии устойчивости организма к гипоксииВ 1983 г. был открыт феномен прерывистой гипоксии, важную роль в котором играет активация митоКАТФ [Murry et al., 1983; Liu et al., 1998]. Феномен прерывистой гипоксии был назван «preconditioning» или «ишемической толерантностью» [Murry et al., 1986; Kitigawa, 1990]. Позднее он получил название «феномен ишемической адаптации» (ФИА) [Yellon et al., 1998; Петрищев и др., 2001], и в настоящее время признан наиболее эффективным способом уменьшения размера инфаркта миокарда [Kloner et al., 1998]. Справедливости ради надо отметить, что адаптация, создаваемая кратковременной гипоксией, была обнаружена ранее [Poupa et al.,1966; Meerson et al., 1973]. Клеточные механизмы ФИА в настоящее время широко изучаются. Считается, что защитный эффект является трехступенчатым и включает в себя последовательные триггерные и конечные эффекторные механизмы, причем наиболее вероятным эффектором является митоКАТФ [Garlid et al., 1997; Yellon et al., 1998; Grover and Garlid., 2000; Петрищев и др., 2001]. ФИА развивается в виде двух волн: первая волна проявляется в течение нескольких часов, затем исчезает и снова появляется в виде второй волны [Yellon et al., 1998], которая длится в течение недели, а иногда и месяца [Carroll et al., 2000]. Установлено, что во второй стадии адаптации к гипоксии также участвует митоКАТФ [Wang et al., 2001]. В развитии толерантности к ишемии мозга [Lacza et al., 2003], и в дистанционной ишемической адаптации сердца к гипоксии при создании коротких эпизодов окклюзии почечной артерии также задействован митоКАТФ [Pell et al., 1998]. Установлено, что специфические модуляторы митоКАТФ (см. таблицу 1) влияют на ФИА. Так активатор митоКАТФ, диазоксид, и неселективный активатор КАТФ каналов, кромакалим, имитируют «preconditioning», увеличивая время сокращения миокарда и способствовуя восстановлению функций изолированных сердец крыс, подвергавшихся общей ишемии миокарда с последующей реперфузией. Кi диазоксида для митоКАТФ (0.8 мкМ в МХ сердца быка) в 1000 раз превышала Кi для цитоКАТФ [Gross et al., 1992]. Кромакалим проявлял аналогичный эффект в тех же концентрациях. Глибенкламид и 5-ГД блокировали действие диазоксида и кромакалима, следовательно, эффект активаторов на ФИА опосредован через КАТФ каналы [Auchampach, 1992; Garlid, 1997; Tsai et al., 1998; Vanden Hoek, 2000]. Активирующе действие диазоксида на митоКАТФ показано также на изолированных МХ сердца крысы, что доказывает роль МХ канала в кардиопротекции [Gross et al., 1992]. Полученные данные подтверждают также результаты исследований, по изучению флуоресценции флавопротеинов на изолированных кардиомиоцитах желудочков сердца кроликов [Liu et al., 1998; Sato et al., 1998]. Однако поскольку было обнаружено, что диазоксид снижает мембранный потенциал на МХ, независимо от активации митоКАТФ канала, его кардиопротекторный эффект вызывает сомнение. Показано, что потенциальный медиатор сигнального пути, ответственного за формирование ишемической адаптации, протеинкиназа С, [Sato et al., 1998; Liu et al., 1994; Sasaki et al., 2000] модулирует активность митоКАТФ. 5-ГД полностью блокирует этот кардиопротекторный эффект, подтверждая участие митоКАТФ в защите миокарда от ишемических повреждений. Следует отметить, что сами по себе глибенкламид и 5-ГД не оказывают никакого влияния [Gross and Fryer, 1999]. Неспособность специфического ингибитора цитоКАТФ, HMR-1883, ингибировать анти-ишемический эффект активаторов митоКАТФ [Fryer et al., 2000], также говорит в пользу участия в защитном механизме МХ канала. Следует отметить, что неселективные активаторы КАТФ каналов, такие как пинацидил, оказывая кардиопротекторное действие, вызывали побочный проаритмический эффект и приводили к существенному понижению давления у подопытных животных [Grover, 1997]. Подобные нежелательные последствия способны свести на нет кардиопротекторное действие данных препаратов. Использование селективных активаторов МХ или цитоплазматического КАТФ каналов позволило бы избежать таких побочных явлений. Можно ожидать, что селективные активаторы цитоКАТФ, такие как HMR-1883, оказывая антиаритмическое действие, не будут негативно влиять на ишемическую адаптацию, вызванную активацией митоКАТФ. Так, показано, что HMR-1883 является потенциальным антифибрилляторным агентом при острой ишемии на модели собаки [Billman et al., 1998]. И наоборот, сайт-специфичные активаторы митоКАТФ не будут оказывать проаритмического действия [Gross and Fryer, 1999]. 2.2.2 Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ Несмотря на то, что роль митоКАТФ в адаптации организма к гипоксии доказана, остается неясным, каким образом открытие канала приводит к защите сердца от ишемических повреждений. Недавно были предложены три различных механизма защиты сердца при гипоксии, которые могут проявляться при открытии митоКАТФ: 1) механизм, при котором усиление входа К+ в МХ ведет к набуханию матрикса, что предупреждает распад АТФ; 2) механизм, основанный на снижении концентрации Са2+ в МХ, что уменьшает вероятность открытия митохондриальной поры и предупреждает гибель кардиомиоцитов; 3) механизм, связанный с изменением уровня активных форм кислорода (АФК), повышенная концентрация которых, с одной стороны, активирует ряд ферментов, а с другой, необратимо повреждает мышечную ткань при гипоксии. Известно, что при набухании МХ усиливается окисление жирных кислот и увеличивается продукция АТФ [Grover, 2000; Halestrap, 1989]. Хотя ранее было продемонстрировано, что набухание МХ является следствием открытия митоКАТФ, вопрос о связи этого эффекта непосредственно с сохранением целостности МХ после ишемии и реперфузии остается открытым [Lim et al., 2002]. Идея о том, что потребление К+ митохондриями и сопутствующее ему набухание этих органелл оказывает кардиопротекторный эффект, подтверждается тем, что активация других митохондриальных К+ каналов, например, Ca2+-активируемого К+ канала, также предупреждает ишемические повреждения [Xu et al., 2002]. Однако, этот эффект не блокируется ингибитором митоКАТФ, что наблюдается при «preconditioning». Следовательно, именно митоКАТФ задействован в формировании феномена «preconditioning». Согласно второй гипотезе [Liu et al., 1999; Murata et al., 2001; Korge et al., 2002; Holmuhamedov et al., 1999], в состоянии аноксии МХ мембрана деполяризуется, скорость электрофоретического входа кальция снижается и, соответственно, уменьшается количество Са2+ в МХ. Это, в свою очередь, предупреждает образование Ca2+-активируемой митохондриальной поры, открытие которой ведет к развитию апоптоза и некроза ткани [Szabo et al., 2004]. Полученные данные согласуются с результатами исследований о предотвращении апоптоза при активации митоКАТФ [Takashi et al., 1999], возможно, путем ингибирования митохондриальной поры [Akao et al., 2003]. Однако, как показано в лаборатории проф. Гарлида, при открытии митоКАТФ снижение потенциала настолько мало [Carreira et al., 2005], что не может существенно отразиться на скорости входа Ca2+ в МХ. Третий механизм, в настоящее время наиболее обсуждаемый, основан на обнаружении изменений уровня АФК, как во время прекондиции [Ozcan et al., 2002], так и в условиях реперфузии [Vanden Hoek et al., 2000]. Стимуляция образования АФК короткими эпизодами гипоксии или при введении активаторов митоКАТФ, предотвращаемая 5-ГД, оказывает защитное действие, ингибируемое антиоксидантами [Forbes et al., 2001; Vanden Hoek et al., 1998]. Этот эффект связан, вероятно, с активацией протеинкиназ, активируемых АФК, что ведет к инициированию целого ряда реакций, приводящих к кардиопротекции [Takashi et al., 1999]. Однако ингибиторы протеинкиназ не всегда устраняют кардиопротекторное действие активатора, как это наблюдалось в случае диазоксида [Krenz et al., 2002]. В противоположность этому АФК, образуемые при реперфузии после продолжительной ишемии, могут быть причиной необратимого клеточного повреждения. Предварительная обработка активаторами митоКАТФ подавляет образование АФК при реперфузии [Pain, et al., 2000; Zweier et al., 1987]. Следовательно, митоКАТФ может способствовать продукции «защитных» АФК при адаптации к гипоксии и уменьшать образование «повреждающих» АФК при реоксигенации. Учитывая все вышесказанное можно сделать вывод, что механизм антиишемического действия активаторов митоКАТФ до сих пор окончательно не выяснен. Кроме того, остается непонятной причина сохранения устойчивости миокарда к гипоксии в течение длительного времени после ишемической адаптации сердца. Именно этот феномен может лежать в основе гипобарической адаптации животных и человека к гипоксии и связан он, вероятно, с экспрессией ряда белков, в том числе и белков, формирующих митоКАТФ [Kuzuya et al., 1993; Marber et al, 1993]. Известно, что синтетические активаторы митоКАТФ оказывают кардиопротекторный эффект. Обнаруженный в нашей лаборатории природный активатор митоКАТФ (УДФ) обладает по сравнению с ними рядом преимуществ, поскольку он не обладает побочными эффектами, которые могут наблюдаться при использовании синтетических препаратов и его концентрацию в клетке легко регулировать. В связи с этим, в работе исследовались анти-ишемическое и антиаритмическое действие уридина и УМФ (предшественников УДФ в клетке [Matsushita et al., 1970]) и опосредованы ли эти эффекты препаратов активацией митоКАТФ. 2.2.3 Феномен прерывистой гипобарической тренировки В любой популяции неинбредных животных существуют особи с различной резистентностью к гипоксии [Березовский, 1978]. Индивидуальные различия в чувствительности к гипоксии и ее переносимости определяются при подъеме животных в барокамере на критическую высоту. Время жизни крайних типов животных (высоко- и низкоустойчивых к гипоксии) на критической высоте различается в 5 и более раз [Березовский, 1978; Чернобаева, Лукьянова, 1989]. Индивидуальные особенности реакции организма на гипоксию играют существенную роль в развитиии, течении и исходе возникающего при этом патологического состояния [Лукьянова и др., 1999; Лукьянова, Коробков, 1981]. Еще в 60-х годах прошлого века было замечено, что в популяциях, живущие на возвышенностях, частота возникновения инфаркта миокарда и смертности от сердечных заболеваний существенно ниже [Hurtado, 1960; Mortimer et al., 1977]. В 1966 г. Поупа с соавторами показали наличие кардиопротекторного эффекта гипобарической гипоксии у крыс с инфарктом миокарда, индуцированным протеренолом [Poupa et al., 1966]. В 1973 Меерсон с соавторами сообщили о том, что симуляция большой высоты 5 минут в день 5 дней в неделю, на 84% сокращает смертность крыс с лигацией коронарной артерии и на 35% размер инфаркта миокарда [Meerson et al., 1973]. Позднее блыло показано, что у крыс, подвергавшихся прерывистой гипоксии, уменьшались желудочковые аритмии, индуцированные ишемией/реперфузией, и лучше сохранялась сократительная функция желудочка [Meerson et al., 1987]. У крыс разных возрастов также увеличивалась устойчивость к аноксии после гипоксической тренировки [McGrath et al., 1973]. В дальнейшем в ряде исследований было показано, у животных, подвергавшихся нескольким предварительным циклам гипоксии, даже в нормобарических условиях, почти на 50% сокращается размер области инфаркта миокарда, вызванного лигированием коронарной артерии [Xi et al., 2002; Cai et al.,2003]. В 2004 Zong с соавторами показали эффективность прерывистой гипоксической тренировки в отношении снижения инфаркта миокарда и аритмии желудочка у собак [Zong et al., 2004]. Это нашло подтверждение и в последующих работах [Downey, 2006]. Интерес исследователей к гипоксической тренировке (ГТ) вызван тем, что она позволяет усилить эффект натренированности, ускоряет акклиматизацию к высоте, предотвращает и лечит некоторые заболевания [Roach., 2001; Levine.,2002; Powell and Garcia., 2000; Boning., 1997; Tin’kov and Aksenov., 2002]. Существуют два типа ГТ: гипобарическая и нормобарическая. В последнем случае животных помещают в камеру, в которую подвется воздух с вдвое сниженной концентрацией кислорода. ГТ включает в себя ~5 минутные эпизоды такой умеренной гипоксии с последующей нормоксией несколько раз в день в течение нескольких недель. Эта процедура не требует сложных приспособлений, а потому может использоваться в клинике [Zong et al., 2004]. Следует подчеркнуть, что Россия является пионеромв этих клинических исследованиях. Кардиопротекторный эффект ГТ аналогичен открытому позднее действию прекондиции [Murry et al., 1983]. Это подтверждается результатами недавних экспериментов, согласно которым, из 46 пациентов с сердчно-сосудистыми заболеваниями, в течение 10 месяцев подвергавшихся ГТ, у 37 инфаркт миокарда не развился [Tin’kov and Aksenov, 2002]. ГТ нормализует и другие патологические состояния [Roach., 2001; Levine.,2002]. Однако, механизм ГТ до сих пор неясен. Колар предложил ряд возможных механизмов для объяснения ее кардиопротекторного действия. 1) Повышенная васкуляризация миокарда и коронарного кровотока, 2) увеличенное содержание гемоглобина в крови и миоглобина в миокарде, 3) оптимизированный энергетический метаболизм, 4) появление специфических нейрогуморальных факторов, простагландинов и стресс белков, 5) увеличенное содержание или активность антиоксидантов, и 6) высвобождение аденозина [Kolar, 1996]. Недавно было высказано предположение о том, что в защитный механизм ГТ вовлечен КАТФ канал [Asemu et al., 1999; Neckar et al., 2002; Zhu et al., 2003]. Так, показано, что у животных, подвергавшихся ГТ, защитный эффект тренировки при ишемии/ реперфузии (30 мин/30 мин) полностью блокировался глибенкламидом и 5-ГД. Следовательно, защитный эффект ГТ опосредуется КАТФ каналами [Zhu et al., 2003]. Усиление же кровотока не является основной причиной защитного действия ГТ [Zong et al., 2004]. Согласно данным Лукьяновой [Лукьянова, 2004], реакция организма на дефицит кислорода является отражением сложного полифункционального ответа клетки, координированного нейрогуморальными механизмами, где в общей иерархии внутриклеточных процессов энергетический обмен выполняет триггерную роль, а нарушение фукции митохондриальных ферментных комплексов являются базисным механизмом любой формы гипоксии. В то же время, вопрос о конечном эффекторе защитного действия ГТ остается открытым. Учитывая данные о роли митоКАТФ канала в кардиопротекции, опосредованной ишемической прекондицией [Garlid, 1997, Liu et al., 1998; Sato et al., 1998], мы предположили, что данный канал может быть вовлечен и в реализацию адаптации при ГТ. Для проверки данного предположения в настоящей работе были исследованы параметры дыхания и АТФ-зависимого калиевого транспорта МХ крыс с различной устойчивостью к ишемии, а также адаптированных к гипоксии. 2.3 Выделение МХ2.3.1 Выделение МХ печени крысы Для выделения МХ использовали самцов крыс альбиносов линии Вистар, массой ~250г. Крыс умерщвляли декапитацией без наркоза. Печень извлекали и помещали в предварительно взвешенную среду выделения (t 0°С). После определения массы и проведения перфузии 0.9% NaCl, печень продавливали через пресс и гомогенизировали в стеклянном гомогенизаторе с тефлоновым пестиком в 8-кратном объеме среды выделения, отнесенном к исходному весу ткани. Среда выделения содержала 250 мМ сахарозы, 10 мМ Tрис-HCl, 0.5 мМ ЭГТА (pH 7,4). Осаждение МХ проводили общепринятым методом дифференциального центрифугирования с модификациями, разработанными в нашей лаборатории (Миронова и др., 1981). К полученному осадку добавляли среду выделения в 0,1-кратном объеме к исходной массе ткани и гомогенизировали. Полученная суспензия МХ, использовавшаяся в дальнейшей работе, содержала 80-100 мг белка/мл. |
|
|||||||||||||||||||||||||||||
|
|||||
Рефераты бесплатно, реферат бесплатно, сочинения, курсовые работы, реферат, доклады, рефераты, рефераты скачать, рефераты на тему, курсовые, дипломы, научные работы и многое другое. |
||
При использовании материалов - ссылка на сайт обязательна. |
||