|
Дипломная работа: Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданиюКонцентрацию белка в МХ определяли по методу Лоури [Лоури, 1951], используя бычий сывороточный альбумин (БСА) в качестве стандарта. В работе использовали самцов крыс линии Вистар, массой ~250г. Предварительные процедуры как в п.п. 1.1. После определения массы сердца, ткань измельчали в растворе, содержащем 300 мМ сахарозы, 10 мМ Hepes, 2 мМ ЭГТА и 10% протеазы в течение 10 минут. Измельченную ткань гомогенизировали стеклянным гомогенизатором с тефлоновым пестиком в 8-кратном объеме среды выделения, отнесенном к исходной массе ткани. Среда выделения содержала 300 мМ сахарозы, 10 мМ Hepes, 2 мМ ЭГТА, 0.1 % бычьего сывороточного альбумина (БСА) (pH 7.4). МХ осаждали дифференциальным центрифугированием [Миронова и др., 1981]. К полученному осадку добавляли среду выделения в 0,1-кратном объеме к исходной массе ткани и гомогенизировали. Полученная суспензия МХ содержала 30-50 мг белка/мл. Концентрацию белка в МХ определяли методом Лоури [Лоури, 1951]. 2.3.2 Выделение и очистка митоКАТФ каналаСолюблизацию КАТФ канала из МХ печени крысы проводили по методу этанольной экстракции, разработанному в нашей лаборатории [Миронова и др., 1981] с некоторыми модификациями. Полученные МХ помещали на 20 мин в гипотонический раствор (концентрация белка составляла 3-4 мг/мл), содержащий 10 мМ Трис-HCl (pH 7,5) при 4° С. Затем экстракт центрифугировали 20 мин при 5500 об/мин. Из полученного осадка митопластов экстрагировали белок. Для этого митопласты разводили 10 мМ Tрис-HCl буфером (рН-7,4) до концентрации 44 мг/мл. К суспензии добавляли 10-кратный водный раствор 66% этанола, охлажденный до -20˚С, и инкубировали при 4˚С в течение 30 минут при постоянном перемешивании. Полученный экстракт центрифугировали при 5500 об/мин в течение 15 минут. Супернатант диализовали против 5 мМ Трис-HCl буфера (рН 7,4) с добавлением 0,05% β-меркаптоэтанола в течение ночи при 4˚С при постоянном перемешивании с одной сменой буфера через 2 часа от начала диализа. Фракцию центрифугировали при 100000 g в ечение 1 часа. Далее проводили ионообменную хроматографическую очистку полученного экстракта. Носитель - ДЕАЕ-целлюлоза (Sigma), объем колонки - 1 см3, диаметр – 0.5 см, h = 5 см. Скорость колонки - 40 мл/ч. Колонка уравновешивалась буфером, содержавшим 50 мМ Трис-HCl, 1 мМ ЭДТА (рН 7,5). Этим же буфером далее элюировали несвязавшиеся с носителем белки. Связавшиеся белки элюировали двумя объемами ступенчатого градиента KCl (50, 100, 150, 200, 250, 300, 500 мМ KCl). После фракционирования белки каждой фракции идентифицировали методом SDS-PAAG электрофореза (10%). Гели окрашивали Кумасси-R250. Исследуемый белок с м.м. 55 кДа элюировался 250 мМ KCl. Глава 3. Изучение энергозависимого входа К+ в МХ методом спектрофотометрии В исследованиях использовались МХ сердца и печени крыс линии Вистар (масса животных ~250г.). Вход ионов калия определяли по скорости набухания МХ в гипотонической среде с KCl. Кинетику набухания регистрировали по изменению оптической плотности суспензии МХ при длине волны 520 нм при постоянном перемешивании и термостатировании при 26°С на спектрофотометре “Uvikon” (Италия). Концентрация МХ белка в ячейке составляла 0.1 мг/мл. Среда инкубации содержала: 50 мМ KCl, 5 мМ HEPES, 5 мМ NaH2PO4, 5мМ янтарной кислоты, 0.5 мМ MgCl2, 0.1 мМ ЭГТА, 5 мкМ цитохрома С, 2 мкM ротенона, 1 мкМ циклоспорина А, рН 7.2. Набухание инициировали добавлением МХ. Скорость набухания рассчитывалась по изменению светорассеяния за единицу времени. 3.1 Изучение ДНФ-индуцированного выхода ионов калия из МХ Проницаемость митохондриальной мембраны оценивали с помощью К+-селективного электрода по содержанию и скорости выхода катиона из деэнергизованных МХ в присутствии разобщителя окислительного фосфорилирования 2,4-динитрофенола (ДНФ). Кинетику выхода калия регистрировали с помощью оригинального электрометрического усилителя, который через контроллер L-153 был соединен с компьютером IBM PC486. Измерения проводились при постоянном перемешивании и термостатировании при 26°С. Концентрация МХ белка в ячейке составляла 1,5-2 мг/мл. Среда инкубации содержала: 0,3 М сахарозы, 3 мМ NaH2PO4, 10 мМ Трис-HCl, pH 7,4. 3.2 Получение и очистка антител к белку с молекулярной массой 55 кДа.3.2.1 Подготовка белка с м.м. 55 кДа: выделение и очистка Выделение и первичная очистка белка проводились методами, описанными в п.п. 2. Белок хранили при температуре -20°С. Фракции, содержавшие белок с м.м. 55 кДа и обладавшие АТФ-ингибируемой К+-селективной активностью, накапливали, затем диализовали против 10 мМ Трис-HCl (рН 7.4) в течение ночи при постоянном перемешивании и температуре 4°С. Обессоленную фракцию концентрировали обратным диализом с использованием полиэтиленгликоля-20000 (ПЭГ) при 4°С. Сконцентрированную фракцию подвергали дополнительной очистке методом нативного электрофореза в полиакриламидном геле (ПААГ). Для электрофореза использовали 10%-ый ПААГ в системе Дэвиса [Davies, 1964]. На геле наблюдалась только одна полоса - белковая зона, в которой определялась АТФ-зависимая К+-транспортирующая активность. Полосу исследуемого белка вырезали из геля, измельчали и элюировали в течение 12 часов при постоянной силе тока (8 мА) и температуре 4°С. Среда элюции содержала 10 мМ Трис-HCl, 38 мМ глицина, рН 8.3. Концентрацию белка в элюате определяли методом Лоури [Лоури, 1951]. Чистоту белка контролировали методом SDS-PAAG электрофореза [Laemmli, 1970]. Выход белка составлял около 30-50 мкг из 100 г ткани печени. К+-транспортирующие свойства выделенного белка и чувствительность к аденозин-5’-трифосфату оценивали при встраивании его в бислойную липидную мембрану. 3.3 Иммунизация и анализ препарата антител Для получения антисыворотки проводили иммунизацию животных электрофоретически чистым АТФ-ингибируемым К+-транспортирующим белком с м.м. 55 кДа (канальная субъединица митоКАТФ), выделенным из печени крысы. В качестве доноров иммунной сыворотки использовали кроликов массой 2.4-2.6 кг. Схема процедуры была подобрана в соответствии с требованиями Декларации совета Европейского Союза 86/609/EEC. За неделю до иммунизации животных дела проводили забор крови из ушной вены; сыворотка в дальнейшем использовали в качестве контроля. Антиген, смешанный с неполным адъювантом Фрейнда (60-100 мкг белка с м.м. 55 кДа растворяли в 300 мкл 0.9 % NaCl и тщательно смешивали с 600 мкл неполного адъюванта Фрейнда) вводили животным подкожно в область лопаток. Гомогенность полученной смеси определяли визуально. Антиген вводили в 10 последовательных инъекций по 100 мкл на небольшом расстоянии друг от друга, чтобы минимизировать болевые ощущения. Через 3 и 6 недель проводили повторные иммунизации по той же схеме, однако для повторных инъекций использовали полный адъювант Фрейнда. Через 7-10 дней после последней инъекции брали кровь из ушной вены (40-50 мл), выдерживали 30 минут в открытом сосуде при комнатной температуре и оставляли на ночь при 4°С. Образовавшийся тромб удаляли центрифугированием (2000 об/мин), а обогащенную антителами сыворотку хранили в замороженном виде при температуре -20°С. 3.3.1 Детекция специфических антител и определение титра Для определения титра полученных антител использовали белок с м.м. 55 кДа, полученный по схеме, описанной в п.п. 5.1.1. Белок в 10 мМ Трис-HCl, рН 7.4, в концентрации 1-2 мкг/мкл сорбировали на полосы нитроцеллюлозной мембраны, в количестве равном числу разведений первичных антител, подсушивали. Затем полоски нитроцеллюлозы инкубировали в ПБС-Твин буфере (0.15 М NaCl, 0.02 М NaH2PO4 × 12 Н2О, рН 7.4, 0.1% Твин-20) в течение 30 минут при комнатной температуре. После этого мембраны в течение часа инкубировали с ПБС-Твин-молоко буфером (ПБС-Твин 20 + 2% сухого молока (Amersham, Германия)) чтобы заблокировать участки, неспецифичного связывания с белком. Буфер для блокирования удаляли и мембраны последовательно инкубировали сначала с первичными специфическими кроличьими антикрысиными антителами, а затем с вторичными антителами (козьи антикроличьи, Sigma), по 1 часу, отмывая мембрану ПБС-Твин-молоко буфере (смена буфера 4-5 раз в течение часа). При этом делали несколько разведений антисыворотки (1:400, 1:800, 1:1600, 1:3200, 1:6400, 1:12800, 1:25600, 1:51200) в ПБС-Твин-молоко буфере. Каждую полоску нитроцеллюлозной мембраны с белком помещали в отдельную емкость и инкубировали с антителами определенного разведения. Разведение вторичных антител – 1:500. Перед окрашиванием полосы мембран тщательно отмывали в ПБС-Твин буфере (4-5 смен буфера в течение 30-40 минут). Окрашивание проводили пероксидазной реакцией в присутствии 0.02% перекиси водорода. Буфер для окрашивания содержал 50 мМ Трис-HCl, рН 7.5. Через 2-5 минут реакцию останавливали, промывая мембрану дистиллированной водой. Затем мембрану высушивали и хранили в темноте. 3.3.2 Вестерн-Блот анализ Данный метод, впервые описанный Товбином и соавторами [Towbin et al., 1979] использовали для идентификации АТФ-зависимого К+-транспортирующего белка с м.м. 55 кДа в различных тканях животных. На первом этапе проводили фракционирование МХ с помощью SDS-PAAG электрофореза [Laemmli, 1979]. Нагрузка составляла 5 мкг суммарного белка. Электрофорез вели при комнатной температуре и постоянном токе (20-30 мА на пластину) в течение 3-4 часов до достижения бромфеноловым синим нижнего фронта геля. После электрофореза половину геля окрашивали Кумасси R-250 или серебрением [Shevchenko et al., 1996], а вторую половину уравновешивали в буфере для блоттинга (150 мМ глицина, 20 мМ Триса, 0.02 % ДСН, 20 % метанол, рН 8.3) в течение 30 минут на качалке при комнатной температуре. Затем гель покрывали нитроцеллюлозной мембраной (Sigma, диаметр пор 0.45 мкм) и помещали между листами влажной фильтровальной бумаги. Систему помещали между двумя пористыми прокладками и зажимали между двумя пластинами из плексигласа. Перенос вели при напряженности 7 В/см2 в течение двух часов при комнатной температуре. После окончания переноса нитроцеллозную мембрану инкубировали в течение 30 минут в ПБС-Твин буфере при комнатной температуре. Дальнейшая процедура такая же, как в п.п. 5.2.1. 3.4 Исследование ДНФ-индуцированного выхода К+ из митохондрий с помощью К+-селективного электрода Функционирование КАТФ-канала в митохондриях оценивали также по инициированной 2,4-динитрофенолом (ДНФ) скорости АТФ- зависимого выхода калия из митохондрий, т.е. создавали условия для работы канала в обратном направлении (Баранова и др, 2000). В среде без субстрата дыхания и калия регистрировали выход калия из митохондрий после добавления разобщителя окислительного фосфорилирования. Выход К+ из митохондрий индуцировали добавлением 50 мкМ 2,4-динитрофенола. Кинетику выхода калия регистрировали с помощью оригинального электрометрического усилителя, который через контроллер L-153 был соединен с компьютером. Измерения производились при постоянном перемешивании. Концентрация митохондриального белка в ячейке составляла 1-1,5 мг/мл. Среда инкубации митохондрий содержала: 170 мМ сахароза, 80 мМ D-маннит, 5 мМ Na2HPO4, 10 мМ Трис-НCl (рН 7.4). 3.5 Реконструкция белка в БЛМ Для реконструкции белка использовали БЛМ, сформированные из смеси 90 % общих липидов мозга быка и 10 % кардиолипина. Плоские бислои формировали методом Мюллера из липидов, растворенных в н-декане (Mueller et al, 1964). Суммарная концентрация липидов в н-декане равна 20 мг/мл. Трансмембранный ток регистрировали при постоянном напряжении на мембране. Проводимость немодифицированной мембраны составляла 1-3 пСм. Раствор белка вводили в буфер, омывающий одну из сторон мембраны. Буфер содержал: 20 мм Tris, 100 мМ KCl (рН 7,4). Регистрацию тока через ионные каналы в мембране проводили при помощи операционного усилителя с высокоомным сопротивлением в цепи обратной связи. Выход усилителя был подключен к компьютеру. Эксперименты проводились при 20-22ºС. 3.6 Иммунноэлектронная микроскопия Ткань печени и сердца фиксировали в 4%-ном растворе параформальдегид/0.05% глутаровый альдегид в PBS – буфере (16.7 мМ Na2HPO4· 12 H2O, 3.3 мМ KH2PO4, 150 мМ NaCl, рН 7,4) в течение 4 ч при 4°С. Обезвоживание в спиртах и пропитку образца смолой LR-White (Sigma, USA) проводили при 4°С. Полимеризацию смолы осуществляли под ультрафиолетом в течение 48 часов при комнатной температуре. Ультратонкие срезы готовили на ультратоме UC6 (Leica, Германия) и помещали на золотые сеточки, покрытые формваровой пленкой и укрепленные углем (Agar). Неспецифическое окрашивание блокировали обработкой раствором, содержащим 3% БСА и 0.5% желатина в течение 1ч. Все дальнейшие процедуры проводили в PBS содержащем 1% БСА и 0.01% тритон Х-100. После каждой инкубации образцы отмывали PBS- буфером, содержащим 0.1% тритон Х-100 и 0.1% глицин и затем в растворе БСА и желатина в течение 20 мин [18]. В качестве первичных антител были использованы полученные нами антитела к митохондриальному К+-транспортирующему белку (в разведении 1:100), инкубацию с которыми проводили в течение ночи при 4°С. После тщательной отмывки сеточки с образцами помещали на каплю вторичных антител, меченных коллоидным золотом с размером гранул 10 нм (Anti-Rabbit IgG, Sigma, USA) и инкубировали в течение 2 часов при комнатной температуре. После отмывки образцы окрашивали уранилацетатом и цитратом свинца и просматривали под электронным микроскопом Tesla BS-500 (Чехословакия). Специфичность метода проверяли заменой первичных антител на буфер. 3.7 MS-MALDI-TOF/TOF- анализ MS-MALDI-TOF/TOF- анализ был выполнен на базе ГУ НИИ биомедицинской химии РАМН им. В.Н. Ореховича. Очищенный митохондриальный К+-транспортирующий белок подвергали ферментативному гидролизу трипсином в денатурирующих условиях в геле. Пептидную смесь из геля экстрагировали ацетонитрил/гидрокарбонат аммонием и затем проводили масс-спектральный анализ на времяпролетном масс-спектрометре Ultraflex (Bruker, Daltonik) в режиме моноизотопической детекции 300-1800 Да. Масс-спектры анализировались через базу данных MSDC и NCBI программой Mascot (http://www.ncbi.nlm.nih.gov). 3.8 Очистка антител к АТФ-зависимому белку с м.м. 55 кДа Для выделения из антисыворотки, содержащей специфические антитела, фракции иммуноглобулинов, использовали методы дробного высаливания, хроматографии и диализа [Антитела. Методы. Изд-во «Мир», 1991, т.1, с.106-107]. Антисыворотку перед высаливанием разводили в 2 раза раствором 0.9% NaCl (рН 7.5). К полученному объему антисыворотки добавляли половинный объем холодного насыщенного раствора сульфата аммония при перемешивании на ледяной бане. Смесь оставляли на 30 минут и затем центрифугировали 20 минут при 5000 g. Осадок растворяли в 0.9% NaCl (рН 7.5). Эту процедуру повторяли 2 раза. Окончательно осадок растворяли в 0.01 М натрий-фосфатном буфере (рН 7.5), содержащем 0.15 М NaCl и диализовали против этого же буфера в течение 18 часов при 4°С. Нерастворившийся осадок удаляли центрифугированием (20000 g, 10 минут). Надосадочную фракцию наносили на колонку (объем 5 см3, диаметр – 1 см, h = 5 см), заполненную ДЭАЭ-целлюлозой (Sigma) и уравновешенную 0.01 М Na-фосфатным буфером (рН 7.5). Элюирование иммунноглобулинов G (IgG) проводили двойным объемом ступенчатого градиента NaCl: 50, 100, 150, 200 мМ. IgG элюировались 50 мМ NaCl. Фракции, содержащие IgG, объединяли и концентрировали с помощью ПЭГ-20000. Полученную фракцию диализовали против 10 мМ Трис-HCl (рН 7.5) в течение ночи при 4°С. Контроль чистоты фракции IgG осуществляли с помощью SDS-PAAG электрофореза [Laemmli, 1979]. 3.9 Очистка антител к АТФ-зависимому белку с м.м. 55 кДа на колонке с иммобилизованным Белком А Перед очисткой иммуноглобулинов в имеющейся сыворотке измеряли концентрацию белка на спектрофотометре Shimadzu UV-2401 РС (Япония) при длине волны 280 нм. Сыворотку разводили 0.1М Na-фосфатным буфером, pH 7.0 до концентрации белка ~2 мг/мл. Разведенную сыворотку, из расчета 20 мг общего белка, наносили на колонку объемом 1 мл, упакованную конъюгированной с белком A (Amersham, Sigma, USA). После нанесения сыворотки колонку промывали тем же буфером до полного отсутствия белка в элюате. Наличие белка в элюате регистрировали при помощи Uvicord S-II LKB (Швеция). IgG элюировали с колонки 0.1М Na-цитратным буфером, pH 3.0. Элюат немедленно титровали 1М Трис-HCl, pH 9.0 до pH 7.0. Затем колонку отмывали 0.1 М Na-фосфатным буфером до pH 7.0. Концентрацию белка в элюате измеряли на спектрофотометре Shimadzu UV-2401 РС (Япония) при длине волны 280 нм. Чистота IgG проверялась при помощи денатурирующего электрофореза в полиакриламидном геле по методу Лэммли [16]. Очищенные IgG разводили глицерином в соотношении 1:1 и хранили при температуре -20ºС. 3.10 Ингибиторный анализ с использованием антител к белкус м.м. 55 кДаАнализ влияния специфических к белку с м.м. 55 кДа антител на параметры функционирования митоКАТФ канала проводили, во-первых, с использованием К+-селективного электрода, определяя скорость ДНФ-индуцированного выхода К+ из МХ и концентрацию ионов К+ в матриксе МХ (см. п.п. 3.2.). Во-вторых, с помощью определения энергозависимого входа К+ в МХ методом спектрофотометрии (см. п.п. 3.1.). При проведении ингибиторного анализа в качестве контроля использовалась преимунная сыворотка, а также сыворотка, содержащая специфические антитела на белок с м.м. 55 кДа, подвергнутая предварительно 5-тиминутному кипячению. Также определялось влияние антител на процесс дыхания МХ. Глава 4. Выделение комплекса цитоплазматических мембран и микросом печени крыс Для выделения комплекса цитоплазматических мембран и микросом печени использовали самцов крыс альбиносов линии Вистар, массой ~250г. Крыс умерщвляли декапитацией без наркоза. Печень извлекали и помещали в предварительно взвешенную среду выделения (t 0°С). После определения массы и проведения перфузии 0.9% NaCl, печень продавливали через пресс и гомогенизировали в стеклянном гомогенизаторе с тефлоновым пестиком в 8-кратном объеме среды выделения, отнесенном к исходному весу ткани. Сначала осаждали МХ дифференциальным центрифугированием, супернатант центрифугировали на 105000 g 1 час, получившийся осадок наносили на градиент (20, 25, 30, 35% сахароза) крутили 105000g 1 час. Происходило разделение образца на две четкие зоны, 25-30% микросомы, 20% мембраны. 4.1 Метод отбора высоко- и низкоустойчивых животных Схема, по которой животные тестировались на устойчивость к гипоксии, была разработана проф. Лукьяновой [Лукьянова и др., 1999; Лукьянова, Коробков, 1981]. В работе использовались самцы крыс линии Вистар массой 250-300 г., которых помещали в барокамеру. Группа высокоустойчивых (ВУ) – животные, которые выдерживали острую гипобарическую гипоксию, соответствующую подъему на высоту 11500 м, в течение 10-15 мин. Группа низкоустойчивых (НУ) животных выдерживала эту высоту только в течение 1-1.5 мин. Глава 5. Результаты и обсуждения 5.1 Параметры функционирования митоКАТФ канала у крыс с различной резистентностью, а также у животных, адаптированных к гипоксии В этом разделе работы исследовались такие показатели, как дыхание МХ, скорость АТФ-зависимого К+ транспорта, количество К+ в МХ, а также чувствительность этого транспорта к АТФ у крыс с различной резистентностью к гипоксии. 5.1.1 Изучение параметров дыхания и окислительного фосфорилирования в МХ печени и сердца крыс с различной резистентностью к гипоксииНесмотря на то, что участие митоКАТФ в защите миокарда и других тканей от ишемических повреждений не вызывает сомнений, механизм позитивного действия активаторов этого канала остается неясным. Для изучения этого вопроса в работе исследовались параметры дыхания и окислительного фосфорилирования в МХ печени и сердца крыс с различной устойчивостью к недостатку кислорода, а также у животных, адаптированных к гипоксии. Изучение параметров дыхания и окислительного фосфорилирования МХ печени и сердца крыс с различной устойчивостью к гипоксии показало, что скорость дыхания МХ во всех метаболических состояниях у высокоустойчивых животных значительно ниже таковой у низкоустойчивых (Рис.5). Следует отметить, что высокоустойчивые животные – это животные, которые выдерживали острую гипобарическую гипоксию, соответствующую подъему на высоту 11500 м, в течение 10-15 мин. Группа низкоустойчивых (НУ) животных выдерживала эту высоту только в течение 1-1.5 мин.
Рисунок 5. Скорость дыхания МХ высоко- и низкоустойчивых к гипоксии НУ – низкоустойчивые к гипоксии животные, ВУ – крысы, высокоустойчивые к гипоксии. Концентрация белка в кювете – 1-2 мг/мл. Среда инкубации: 5 мМ Tris-HCl, 200 мМ сахарозы, 50 мМ KCl, 5 мМ NaH2PO4, 3 мкМ ротенона, рН 7.2. Эксперименты проводились в закрытой ячейке при постоянном перемешивании и термостатировании при температуре 26°С. При этом измерение дыхательного контроля времени фосфорилирования АДФ показало, что окислительное фосфорилирование, то есть синтез АТФ происходит более эффективно у высокорезистентных крыс (Рис.6).
Рисунок 6. Дыхательный контроль и время фосфорилирования МХ крыс с различной устойчивостью к гипоксии ВУ – высокоустойчивые к гипоксии крысы, НУ – низкоустойчивые к гипоксии животные. Условия как на рис. 7. Это свидетельствует об исходно меньшей экономичности процесса синтеза АТФ у низкоустойчивых животных. Таким образом, гипоксия, т.е. увеличение функциональной нагрузки на дыхательную цепь МХ при подъеме животных на высоту или при подаче воздуха со сниженной концентрацией кислорода, может привести к истощению резервных возможностей дыхательной цепи, что не происходит у высокоустойчивых животных. Адаптация низкоустойчивых крыс интервальной нормобарической гипоксией по методу, предложенному в Hypoxia Medical Academy (США), проявляется у них в сопряжении дыхания, что, в свою очередь, выражается в снижении скорости дыхания, увеличении дыхательного контроля и сокращении времени, необходимого для фосфорилирования АТФ (Рис.9).
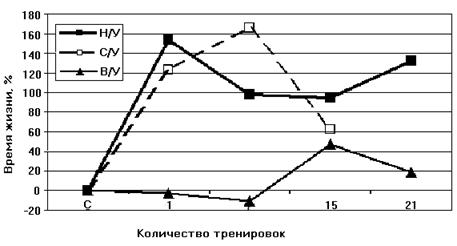
Как видно из рис. 7, при адаптации к недостатку кислорода животных низкоустойчивых к гипоксии, скорость дыхания МХ у них уменьшается. Кроме того, у адаптированных крыс сокращается время фосфорилирования и увеличивается значение дыхательного контроля (Рис.9 Б). Эти данные свидетельствуют о возрастании степени сопряженности дыхательной цепи МХ крыс низкоустойчивых к килородному голоданию при гипоксической тренировке. Следовательно, адаптация приводит к переходу крыс из состояния низкой устойчивости в состояние повышеной устойчивости к гипоксии. Согласно данным Лукьяновой [Лукьянова, 2004] при адаптации крыс гипоксической тренировкой время жизни животного при подъеме на высоту, характеризующее общую неспецифическую резистентность крыс, у НУ животных увеличивается уже после первой тренировки на 120-160% (контроль принят за 0). У НУ время жизни в последующие 20 дней превышает исходную резистентность в 2 раза. У ВУ изменения во времени жизни вообще отсутствуют в первые 7 дней, после чего наблюдается небольшое увеличение, максимум до 40%. В работе делается вывод, что при адаптации гипоксической тренировкой наблюдается экономизация энергетических процессов МХ (Рис. 8).
Рисунок 8. Динамика изменения устойчивости к кислородной недостаточности при гипоксической тренировке животных с различной устойчивостью к гипоксии Н/У – низкоустойчивые крысы, В/У – высокоустойчивые, С/У – животные со средней степенью устойчивости (выдерживающие подъем на высоту в 11500 м в течение 7 минут) [Лукьянова, 2004]. В случае, когда такая оптимизация уже существует, как у высокорезистентных животных, тренировки не приводят к увеличению времени жизни крыс в условиях гипоксии [Лукьянова, 2004]. Поэтому эксперименты по адаптации животных мы проводили на низкоустойчивых к гипоксии крысах. |
|
|||||||||||||||||||||||||||||
|
|||||
Рефераты бесплатно, реферат бесплатно, сочинения, курсовые работы, реферат, доклады, рефераты, рефераты скачать, рефераты на тему, курсовые, дипломы, научные работы и многое другое. |
||
При использовании материалов - ссылка на сайт обязательна. |
||

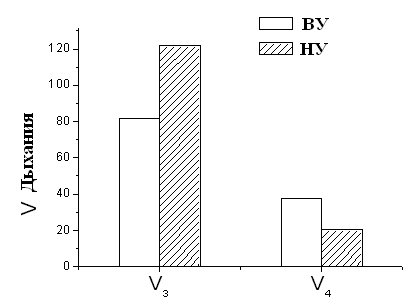
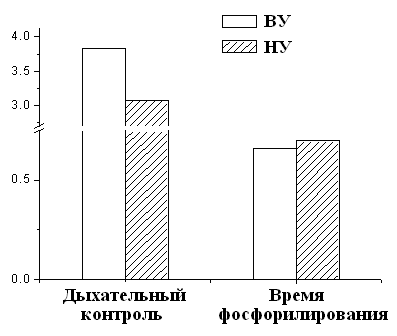
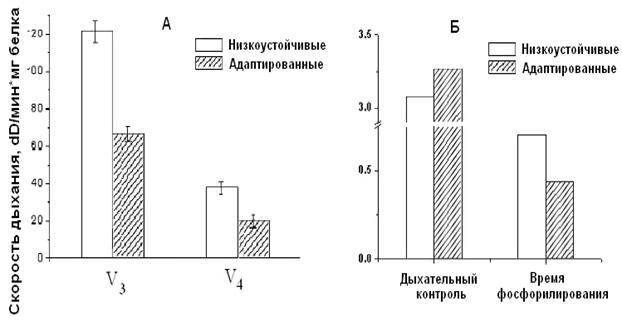 Рисунок 7. Скорость
дыхания (А) и время фосфорилирования (Б) МХ сердца крыс низкоустойчивых и
адаптированных к гипоксии
Рисунок 7. Скорость
дыхания (А) и время фосфорилирования (Б) МХ сердца крыс низкоустойчивых и
адаптированных к гипоксии